Bok om teistisk evolusjon siterer avkreftede ikoner
Av Casey Luskin 19. mai 2026, Oversatt herfra. {kursiv og understreking ved oversetter.}
 I et tidligere innlegg bemerket jeg at den teistiske evolusjonistiske fysikeren Stephen Barr har en artikkel på First Things som overdriver den antatte "gradvise" naturen til menneskets opprinnelse. En av Barrs hovedkilder som han roser i artikkelen sin, er en bok av en katolsk teistisk evolusjonist, Daniel Kuebler, professor i biologi ved Franciscan University of Steubenville. Barr kaller Kueblers bok, Darwin and Doctrine: The Compatibility of Evolution and Catholicism, "en utmerket ny bok" som forklarer "vitenskapen om evolusjon, og som på en nyttig måte rydder opp i vanlige misforståelser underveis".
I et tidligere innlegg bemerket jeg at den teistiske evolusjonistiske fysikeren Stephen Barr har en artikkel på First Things som overdriver den antatte "gradvise" naturen til menneskets opprinnelse. En av Barrs hovedkilder som han roser i artikkelen sin, er en bok av en katolsk teistisk evolusjonist, Daniel Kuebler, professor i biologi ved Franciscan University of Steubenville. Barr kaller Kueblers bok, Darwin and Doctrine: The Compatibility of Evolution and Catholicism, "en utmerket ny bok" som forklarer "vitenskapen om evolusjon, og som på en nyttig måte rydder opp i vanlige misforståelser underveis".
Bilde 1. DNA som digitale data
Kueblers bok har mange teologiske aspekter som jeg ikke tør å kommentere siden jeg ikke er katolikk. Jeg kan imidlertid si at mange av de vitenskapelige argumentene som Kuebler bruker for å argumentere for evolusjon, er gammeldagse argumenter som har blitt behandlet og tilbakevist i årevis. Jeg vil gjennomgå mange av disse problemstillingene nedenfor - men fordi vi har tatt opp så mange av dem flere ganger før, vil mye av svaret mitt ganske enkelt være å sitere fra eller lenke til tidligere artikler. Og vær oppmerksom på at selv om jeg vil vise at mye av Kueblers argument er feilaktig, er det ingenting personlig. Etter alt å dømme er professor Kuebler en anstendig person som søker sannhet, slik som resten av oss. Men hvis du skal akseptere evolusjonsteorien, gjør det i det minste av gode grunner.
Den genetiske kodens universalitet
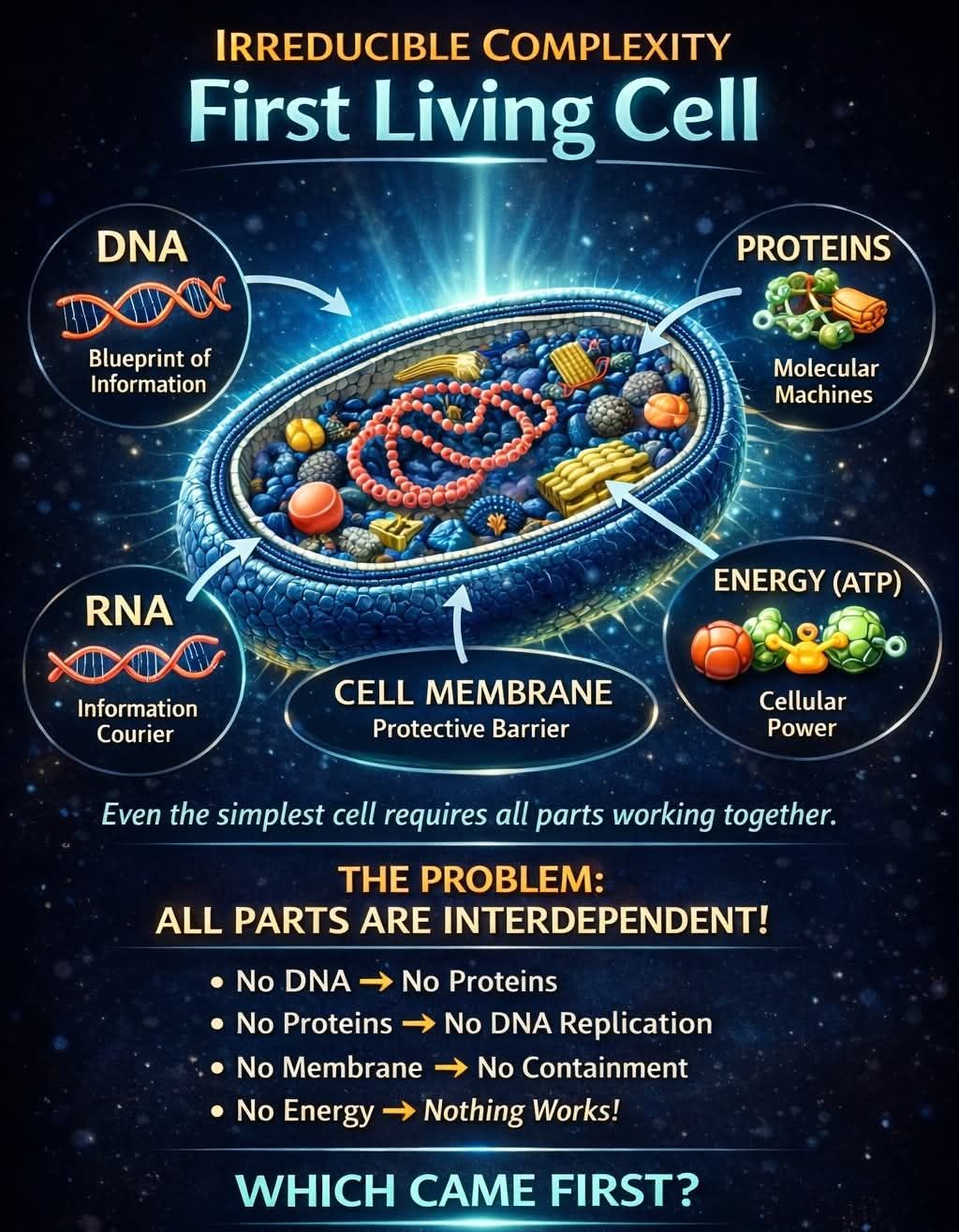 Professor Kuebler bruker et klassisk argument for felles avstamning, og viser til den genetiske kodens universalitet, som han kaller "sannsynligvis det beste beviset for universell felles avstamning" (s. 107). Her er hva Kuebler skriver:
Professor Kuebler bruker et klassisk argument for felles avstamning, og viser til den genetiske kodens universalitet, som han kaller "sannsynligvis det beste beviset for universell felles avstamning" (s. 107). Her er hva Kuebler skriver:
Det finnes en god del indisier for at alle livsformer faktisk er i slekt via felles avstamning. Det mest "universelle" beviset er det faktum at alle livsformer bruker i utgangspunktet den samme genetiske koden. (s. 106)
Problemet med dette argumentet er at det ikke er sant. Som NCBIs nettsted forklarer, finnes det dusinvis av varianter av den genetiske koden. Kuebler gir knapt det minste hint om at disse variantgenetiske kodene eksisterer, og begraver i en fotnote erkjennelsen av at "Det finnes noen få organismer som har små modifikasjoner av den genetiske koden." (s. 107)
Men selvfølgelig er det sant at de aller fleste organismer bruker den samme genetiske koden. Kuebler sier at det ikke finnes noen god grunn til dette, og skriver:
Selv om den universelle genetiske koden er ganske robust, finnes det en rekke andre mulige genetiske koder som ville fungert like bra som den som finnes i så godt som alle celler; faktisk kan noen alternative versjoner fungere enda bedre. Akkurat som det finnes mange forskjellige menneskelige språk, hvorav de fleste fungerer ganske bra, kunne det ha vært mange forskjellige "språk" av den genetiske koden. Det faktum at nesten alle organismer deler dette bestemte språket, selv om det er mange som ville ha passet, antyder at koden oppsto i en felles forfader og ble arvet av alle påfølgende livsformer via felles avstamning. (s. 107)
Bilde 2. I levende celler henger alt sammen (Kr. O. Grasso)
Med andre ord, siden det ikke finnes noen god grunn til at alle disse organismene skal bruke den genetiske koden vi bruker, må forklaringen være felles avstamning. Det er et klassisk argument at ineffektiv og klønete evolusjon gjorde det.
Men det finnes svært gode grunner til å bruke den genetiske koden som finnes i de fleste livsformer. Som Jonathan McLatchie forklarer: Denne ordningen er langt fra vilkårlig. Den genetiske koden som finnes i naturen er faktisk utsøkt innstilt for å beskytte cellen mot de skadelige effektene av substitusjons- mutasjoner. Systemet er så briljant satt opp at kodoner som avviker med bare én base enten spesifiserer den samme aminosyren, eller en aminosyre som er medlem av en beslektet kjemisk gruppe. Med andre ord er strukturen til den genetiske koden satt opp for å redusere effektene av feil som kan oppstå under translasjon (som kan oppstå når et kodon oversettes av et nesten komplementært antikodon).
 Det finnes en rekke andre spesielle egenskaper ved vår genetiske kode som McLatchie dokumenterer her og her.
Det finnes en rekke andre spesielle egenskaper ved vår genetiske kode som McLatchie dokumenterer her og her.
"Livets løsning"
Forresten, Kueblers sitering av påstanden om at "noen alternative versjoner" av den genetiske koden "kan fungere enda bedre" er fra kapittel 1 og 2 i Simon Conway Morris’ bok Life’s Solution. Det er en flott bok, men siteringen hans, som refererer til arbeidet til Steven Freeland og Laurence Hurst, viser faktisk hvor effektiv standard genetisk kode er:
De skriver: "den naturlige genetiske koden viser oppsiktsvekkende bevis på optimalisering, to størrelsesordener høyere enn det som tidligere har blitt antydet. Selv om den presise kvantifiseringen som brukes her kan stilles spørsmål ved, virker det samlede resultatet ganske klart: under vår modell, av 1 million tilfeldige variantkoder produsert, var bare 1 bedre ... enn den naturlige koden - vår genetiske kode er bokstavelig talt "1 av en million"." (s. 17)
Conway Morris antyder at "det er også en følelse av at gitt en verden av DNA og aminosyrer, er kanskje den genetiske koden vi kjenner mer eller mindre et uunngåelig resultat" (s. 18). Jeg ser ingen bevis i Conway Morris’ bok for at det finnes andre koder som er bemerkelsesverdig bedre enn den universelle koden.
Når det gjelder variantkodene, fant Winston Ewert i sin artikkel "On the Origin of the Codes: The Character and Distribution of Variant Genetic Codes is Better Explained by Common Design than Evolutionary Theory" at variantene ofte korrelerer med organismens livsstil, noe som tyder på at også disse forklares av intelligent design:
Bilde 3. Bok om teistisk evolusjon (Otangelo Grasso)
Den nesten universelle genetiske koden siteres ofte som bevis for universell felles aner. På den annen side peker kritikere av universell felles aner ofte på unntak fra den universelle koden som bevis mot den. Det har imidlertid aldri vært en omfattende undersøkelse av karakteren og fordelingen av variantgenetiske koder og deres implikasjoner for debatten om universell felles aner. Denne artikkelen utvikler et rammeverk for å forstå koder innenfor et felles designrammeverk, basert på premisset om at noen genetiske kodevarianter er designet og andre er et resultat av mutasjoner i oversettelsesmaskineriet. Vi fant at disse to kildene til variantkoder kan skilles fra hverandre ved å vurdere organismens livsstil, taksonomisk rang, evolusjonær gjennomførbarhet, kodon-sjeldenhet og distribusjonskompleksitet. Disse ulike tilnærmingene til å skille kodene gir svært korrelerte resultater, noe som viser imponerende forklaringskraft for vårt rammeverk. I motsetning til dette finner vi at evolusjonsteorien har vanskeligheter med å forklare karakteren og fordelingen av variantgenetiske koder.
I den grad koden brukes mye om igjen i forskjellige organismer, eller varierer i forskjellige organismer, kan den dermed forklares med intelligent design.
Gjenbruk av biomolekyler og fylogenetiske trær
 Kuebler tilbyr et annet argument for universell felles avstamning - nemlig gjenbruk av de samme biomolekylene og vår evne til å konstruere fylogenetiske trær. Han skriver:
Kuebler tilbyr et annet argument for universell felles avstamning - nemlig gjenbruk av de samme biomolekylene og vår evne til å konstruere fylogenetiske trær. Han skriver:
Selv om den genetiske koden sannsynligvis er det beste beviset for universell felles avstamning, finnes det også andre bevis. Alle celler bruker dobbelttrådet DNA-molekyler for å lagre informasjon, de er alle bundet av lignende typer lipidmembraner, og de deler mange av de samme grunnleggende veiene for å produsere og bruke energi i form av ATP-molekyler og for å utføre andre viktige livsopprettholdende reaksjoner.
Bilde 4. ID ingens vitenskapsstopper
Ironisk nok, i kapittelet mitt "Universal Common Descent: A Comprehensive Critique" i bindet Theistic Evolution: A Scientific, Philosophical, and Theological Critique, både forutså og besvarte jeg denne samme argumentasjonen:
Kanskje det vanligste argumentet for universell felles avstamning som studenter møter i biologibøker på universitetsnivå, er universaliteten til den genetiske koden - påstanden om at alt liv bruker nukleotidtriplettene til å kode for de samme aminosyrene. Den genetiske koden er imidlertid ikke universell; mange varianter i den genetiske koden er kjent blant ulike organismer.
 Hvis universaliteten til den genetiske koden gir bevis for universell felles aner, teller dens ikke-universalitet som bevis imot? Uansett svar, til tross for variantene bruker de aller fleste organismer den samme "standardkoden", og alle livsformer bruker lignende typer biomolekyler, som nukleotider og proteiner. Er slike utbredte biomolekylære likheter bevis for felles aner? En artikkel fra 2010 i Nature, "En formell test av teorien om universell felles aner", argumenterte ja:
Hvis universaliteten til den genetiske koden gir bevis for universell felles aner, teller dens ikke-universalitet som bevis imot? Uansett svar, til tross for variantene bruker de aller fleste organismer den samme "standardkoden", og alle livsformer bruker lignende typer biomolekyler, som nukleotider og proteiner. Er slike utbredte biomolekylære likheter bevis for felles aner? En artikkel fra 2010 i Nature, "En formell test av teorien om universell felles aner", argumenterte ja:
"[D]et 'universelle' i universell felles aner støttes primært av to ytterligere bevislinjer: ulike viktige fellestrekk på molekylært nivå (inkludert grunnleggende biologiske polymerer, genetisk materiale fra nukleinsyrer, L-aminosyrer og kjernemetabolisme) og den nesten universelle genetiske koden."
Bilde 5. Ribosomet IRK (Ikke-reduserbar-kompleksitet)
Artikkelens forfatter, evolusjonsbiokjemikeren Douglas Theobald, konkluderte med at universell felles aner er den "beste" forklaringen på disse biomolekylære likhetene. Men "best" sammenlignet med hva? Theobald testet universelle felles aner mot den svært usannsynlige hypotesen om at levende organismer uavhengig utviklet de samme bio-molekylene og sekvensene ved ren "tilfeldighet". Universell felles aner virket bare overbevisende fordi den ble sammenlignet med en absurd nullhypotese. Som kritikere som skrev i Biology Direct observerte:
"Cogniscenti grøss da de så Theobald-artikkelen, vel vitende om at 'den er triviell'. Den er triviell fordi stråmannen som Theobald angriper i en tekst som i stor grad er formulert i innviklet juridisk språk, er at betydelig sekvenslikhet kan oppstå ved tilfeldighet i motsetning til avstamning med modifikasjon."
Riktignok er universell felles aner en mulig forklaring på genetiske likheter - men finnes det andre mulige forklaringer? Ja, det er sant. Intelligente agenter bruker ofte de samme delene i forskjellige design for å oppfylle funksjonelle krav, for eksempel gjenbruk av hjul på biler og fly, eller gjenbruk av viktige datakoder i forskjellige versjoner av Microsoft Windows. Som Paul Nelson og Jonathan
Wells observerer:
"En intelligent årsak kan gjenbruke eller omplassere den samme modulen i forskjellige systemer, uten at det nødvendigvis er noen materiell eller fysisk forbindelse mellom disse systemene. Enda enklere kan intelligente årsaker generere identiske mønstre uavhengig av hverandre. .. Hvis vi antar at en intelligent designer konstruerte organismer ved hjelp av et felles sett med polyfunksjonelle genetiske moduler - akkurat som menneskelige designere for eksempel kan bruke den samme transistoren eller kondensatoren i en bilradio eller en datamaskin, enheter som ikke er "homologe" som artefakter - kan vi forklare hvorfor vi finner de "samme" genene uttrykt i utviklingen av det som er svært forskjellige organismer."
 Dermed er felles design - den bevisste gjenbruken av en felles blåkopi eller komponenter - en levedyktig forklaring på de utbredte funksjonelle likhetene mellom biomolekylene som finnes i forskjellige typer organismer. Universell felles opprinnelse er ikke den eneste levedyktige forklaringen.
Dermed er felles design - den bevisste gjenbruken av en felles blåkopi eller komponenter - en levedyktig forklaring på de utbredte funksjonelle likhetene mellom biomolekylene som finnes i forskjellige typer organismer. Universell felles opprinnelse er ikke den eneste levedyktige forklaringen.
Bilde 6. Fellesa avstamning -forutsetning eller konklusjon?
(I motsetning til Theobalds argumenter for universell felles opphav, er ikke alle grunnleggende biomolekyler universelle blant organismer. Som en artikkel fant, er "flere kjernekomponenter i det bakterielle [DNA]-replikasjonsmaskineriet urelaterte eller bare fjernt beslektet med de funksjonelt ekvivalente komponentene i det arkeologiske/eukaryotiske replikasjonsapparatet", noe som førte til at de antydet at "DNA-replikasjon sannsynligvis utviklet seg uavhengig i de bakterielle og arkeologiske/eukaryotiske avstamningene". Enda mer slående er det at en annen artikkel sammenlignet genomene til 1000 forskjellige prokaryote organismer og fant at "av de 1000 tilgjengelige genomene er ikke et eneste protein bevart på tvers av alle genomene.")
ID-teoretikere har også erkjent at effektiv utforming av biologiske Molekyler og signalveier kan tjene som en bedre forklaring, sammenlignet med felles avstamning, på hvorfor de brukes på nytt.
"Omvendt systemteknikk" (Reversed engineering)
Glykolyse er en viktig og universell metabolsk vei som utfører viktige funksjoner som å produsere energi eller bidra til å syntetisere livets byggesteiner. Biokjemiker Emily Reeves og ingeniør Gerald Fudge publiserte en fagfellevurdert studie som brukte ID-basert "omvendt systemteknikk" for å forstå hvorfor glykolyse fungerer som den gjør:
-Biologiske systemer viser egenskaper som tradisjonelt er assosiert med god ovenfra-og-ned kravdrevet systemteknikkpraksis, som modularitet, optimalitet, robusthet, felles protokoller og gjenbruk av design.
Bilde 7. Ingen sikre bevis for felles avstamning i følge E. Koonin
 De konkluderte med at selv om "den nesten ensartetheten av sentral metabolisme gjennom livet tradisjonelt har blitt tilskrevet utelukkende universell felles avstamning", kan likevel denne "ensartetheten forventes" "fra et systemteknisk perspektiv" på grunn av dens funksjonelle eleganse: Glykolyseveier "maksimerer termodynamisk effektivitet" og bruker "resirkulerbare avfallsprodukter" som "forenkler vedlikehold av økosystemhomeostase".
De konkluderte med at selv om "den nesten ensartetheten av sentral metabolisme gjennom livet tradisjonelt har blitt tilskrevet utelukkende universell felles avstamning", kan likevel denne "ensartetheten forventes" "fra et systemteknisk perspektiv" på grunn av dens funksjonelle eleganse: Glykolyseveier "maksimerer termodynamisk effektivitet" og bruker "resirkulerbare avfallsprodukter" som "forenkler vedlikehold av økosystemhomeostase".
Med andre ord, jo mer vi ser på biologi, desto mer innser vi at det er gode grunner til at ting er utformet som de er. Evolusjonister som Kuebler unnlater å undersøke disse gode designprinsippene som nesten alltid er innebygd i biologien. I stedet antar de ganske enkelt at felles opphav er årsaken til gjenbruk, og går glipp av spennende undersøkelser basert på ID-teknikk.
...........
Dårlige synteni-argumenter hevder "ingen funksjonell grunn" for genomiske arrangementer
Av Casey Luskin 20. mai 2026. Oversatt herfra
I et tidligere innlegg bemerket jeg at en relativt ny bok av Daniel Kuebler, professor i biologi ved Franciscan University of Steubenville, siterer noen vanlige, men tilbakeviste evolusjonære argumenter. Mange av disse behandlet jeg i kapittelet mitt "Universal Common Descent: A Comprehensive Critique" i boken Theistic Evolution: A Scientific, Philosophical, and Theological Critique fra 2017. Som kapittelet mitt sier, hevder folk ofte at vår evne til å konstruere fylogenetiske trær gir bevis for felles avstamning. Og Kuebler argumenterer faktisk for dette:
"Ytterligere overbevisende støtte for universell felles avstamning kan finnes ved å se på bevisene for felles avstamning på de ulike grenene av det evolusjonære treet. Selv om det ikke fullt ut ville etablere universell felles avstamning, ville bevis for at alle primater er i slekt med felles avstamning eller at alle pattedyr er forent med felles avstamning være i samsvar med og gi sterk støtte for universell felles avstamning." (s. 107-108)
Likevel gir forsøk på å rekonstruere fylogenetiske trær ved hjelp av molekylære eller morfologiske trekk ofte notorisk inkonsistente resultater. Jeg har forklart dette utallige ganger, som her, her og her. Interessant nok dveler ikke Kuebler ved konsistensen av fylogenetiske trær. I stedet fokuserer han på eksistensen av delte "synteniblokker". Her er hva han sier:
"Et spesielt sterkt bevis som støtter denne typen lokal felles avstamning, er eksistensen av homologe synteniblokker, som er lignende regioner av DNA som er bevart i forskjellige arter. Man kan tenke på disse synteniblokkene som genompuslespill som eksisterte i en felles forfader, men som er satt sammen på forskjellige måter i deres etterkommere på grunn av ulike storskala genomarrangementer som har skjedd over tid."
 For å hoppe til poenget, sier han at
For å hoppe til poenget, sier han at
"arter så fjernt beslektet som mennesker og mus deler et stort utvalg av synteniblokker. Gitt at det ikke ser ut til å være noen funksjonell grunn til at genene som finnes i disse synteniblokkene skal være i en lignende rekkefølge i to forskjellige arter (sannsynligheten for at dette skjer ved en tilfeldighet er forsvinnende liten), er den mest sparsommelige forklaringen at de ble arvet i den spesifikke rekkefølgen fra en delt felles forfaderart." (s. 108-109)
Bilde 8. Mus -felles gener med menneske
Svært utdatert vitenskap
Problemet med dette argumentet er at det er basert på svært utdatert vitenskap. Faktisk er det sjokkerende for meg at en biolog på 2020-tallet ville hevde at det ikke finnes noen "funksjonell grunn" til hvordan gener er organisert på kromosomer. Mye av det vi har lært om genomikk viser at det finnes viktige grunner, sentrale i genomfunksjonen, til hvorfor kromosomer er strukturert slik de er. Jeg forklarte det her:
"Synteni refererer til storskala likheter mellom genomer. [Noen teistiske evolusjonister] hevder at storskala 3D-funksjoner i genomet ikke har noen funksjonell betydning. Dette er rett og slett feil. Mange artikler viser at den storskala 3D-organiseringen av genomet er avgjørende for genomfunksjonen. Etter hvert som revolusjonen innen epigenetikk har tatt fotfeste, vet molekylærbiologer nå at strukturen til kromosomer, og deres 3D-arrangement(er) i en celle, er viktige deler av genomisk regulering."
En rekke artikler viser at kromosomstruktur og genrekkefølge kan være funksjonelt viktig - inkludert følgende:
*Jachowicz et al., "Heterochromatin establishment at pericentromeres depends on nuclear position," (Heterokromatin-etablering ved pericentromerer avhenger av kjerneposisjon) Genes & Development, 27: 2427-2432 (2013);
*Verdaasdonk et al., "Centromere Tethering Confines Chromosome Domains," Molecular Cell, 52: 1-13 (December 26, 2013);
*Filion et al., "Systematic Protein Location Mapping Reveals Five Principal Chromatin Types in Drosophila Cells," (Systematisk proteinlokasjonskartlegging avslører fem viktigste kromatintyper i Drosophila-celler) Cell, 143: 212-224 (October 15, 2010);
*Giacomo Cavalli, "From Linear Genes to Epigenetic Inheritance of Three-dimensional Epigenomes," (Fra lineære gener til epigenetisk arv av tredimensjonale epigenomer) Journal of Molecular Biology (2011);
 *Justin M. O’Sullivan, "Chromosome Organizaton in Simple and Complex Unicellular Organisms," (Kromosomorganisering i enkle og komplekse encellede organismer) Current Issues in Molecular Biology, 13: 37-42 (2011);
*Justin M. O’Sullivan, "Chromosome Organizaton in Simple and Complex Unicellular Organisms," (Kromosomorganisering i enkle og komplekse encellede organismer) Current Issues in Molecular Biology, 13: 37-42 (2011);
Bilde 9. Menneskelemmer er finstemte og optimale
*Dirar Homouz and Andrzej S. Kudlicki, "The 3D Organization of the Yeast Genome Correlates with Co-Expression and Reflects Functional Relations between Genes," (3D-organiseringen av gjærgenomet korrelerer med samekspresjon og reflekterer funksjonelle forhold mellom gener) PLoS One, 8: e54699 (January, 2013);
*Stephen A. Hoang and Stefan Bekiranov, "The Network Architecture of the Saccharomyces cerevisiae Genome," PLoS One, 8: e81972 (December, 2013).
*Petkov PM, Graber JH, Churchill GA, DiPetrillo K, King BL, Paigen K. Evidence of a large-scale functional organization of mammalian chromosomes. (Bevis for en storskala funksjonell organisering av pattedyrkromosomer) PLoS Genet. 2005 Sep;1(3):e33.
*Graber JH, Churchill GA, Dipetrillo KJ, King BL, Petkov PM, Paigen K. Patterns and mechanisms of genome organization in the mouse. (Mønstre og mekanismer for genomorganisering hos mus) J Exp Zool A Comp Exp Biol. 2006 Sep 1;305(9):683-8.
 *Spellman PT, Rubin GM. Evidence for large domains of similarly expressed genes in the Drosophila genome. (Bevis for store domener av lignende uttrykte gener i Drosophila-genomet) J Biol. 2002;1(1):5. doi: 10.1186/1475-4924-1-5.
*Spellman PT, Rubin GM. Evidence for large domains of similarly expressed genes in the Drosophila genome. (Bevis for store domener av lignende uttrykte gener i Drosophila-genomet) J Biol. 2002;1(1):5. doi: 10.1186/1475-4924-1-5.
*Boutanaev AM, Kalmykova AI, Shevelyov YY, Nurminsky DI. Large clusters of co-expressed genes in the Drosophila genome. Nature. (Store klynger av samuttrykte gener i Drosophila-genomet. = 2 Dec 12;420(6916):666-9.
*de Wit E, Braunschweig U, Greil F, Bussemaker HJ, van Steensel B. Global chromatin domain organization of the Drosophila genome. (Global kromatindomeneorganisering av Drosophila-genomet) Genet. 2008 Mar 28;4(3):e1000045. doi: 10.1371/journal.pgen.1000045.
Bilde 10. Star variasjon i kromosom-antall
Molekylbiologer vet nå at kromosomenes struktur og deres tredimensjonale arrangement(er) i en celle er viktige deler av genomisk regulering. Ideen om at "det ikke ser ut til å være noen funksjonell grunn" til at gener og kromosomer skal være ordnet og strukturert på bestemte måter, tilbakevises av det enorme antallet oppdagelser vi gjør innen genomikk.
Man kan bli fristet til å føle seg sjokkert over at så sterke bevis i hovedsak blir ignorert. Likevel er det kanskje ikke så sjokkerende når man innser at evolusjonære argumenter har en veldig lang og skitten historie med feilaktig å hevde at noen funksjoner ikke er funksjonelle, og at de derfor er bevis for evolusjon.

Bilde 11. Casey Luskin
Casey Luskin er geolog og advokat med høyere grad i vitenskap og jus, som gir ham ekspertise i både de vitenskapelige og juridiske dimensjonene av debatten om evolusjon. Han fikk sin doktorgrad i geologi fra University of Johannesburg, og BS- og MS-grader i geovitenskap fra University of California, San Diego, hvor han utstrakt studerte evolusjon, både på hoved- og lavere nivå. Hans jusgrad er fra University of San Diego, hvor han fokuserte studiene på første Amendment, utdanningslov og miljørett.
Oversettelse, tillatt av Discovery Institute, v. Asbjørn E. Lund
---------